Razne. B. Pasmine. V. Strain. G. Race.
8. Navedite faktor koji je glavni u stvaranju novih sorti i rasa..
A. Nasljednost. B. Varijabilnost. B. Veštačka selekcija. D. Prirodna selekcija.
9. Imenujte odnos polova tokom polne reprodukcije.
A. 2: 2. B. 1: 1. C. 3: 1. D. 4: 1.
10. Navedite fazu ontogeneze kojom se završava formiranje neuralne cijevi kod sisara.
A. Blastula. B. Gastrula. B. neurulae. G. Embrion.
11. Zabilježite osobinu koja nije karakteristična za mutaciju.
ODGOVOR: Nastaju iznenada, naglo. B. Imaju adaptivni značaj.
B. Naslijeđeno. D. Nisu rasprostranjeni.
12. Imenujte polne hromozome pilića.
A. xx. B. Hu. V. ho. D. Odsutan.
ekologija se počela razvijati kao samostalna nauka 5. pravac kretanja prirodne selekcije diktira 6. Faktori okoline koji utiču na organizam 7. Grupa faktora sredine uzrokovanih uticajem živih organizama 8. Grupa faktora sredine uzrokovanih uticajem uticaj živih organizama 9. Grupa faktora sredine uzrokovanih uticajem nežive prirode 10. Faktor nežive prirode koji daje podsticaj sezonskim promenama u životu biljaka i životinja. 11. sposobnost živih organizama da imaju sopstvene biološke ritmove u zavisnosti od dužine dnevnog svetla 12. Najznačajniji faktor za preživljavanje 13. Među faktorima su svetlost, hemijski sastav vazduha, vode i zemljišta, atmosferski pritisak i temperatura 14. izgradnja željeznica, oranje zemlje, stvaranje rudnika odnosi se na 15. Predacija ili simbioza se odnosi na faktore 16. dugovječne biljke 17. kratkovječne biljke 18. biljke tundre uključuju 19. biljke polupustinjske, stepske i pustinjske uključuju 20. Karakteristični pokazatelj populacije. 21. Ukupnost svih vrsta živih organizama koji naseljavaju određenu teritoriju i međusobno djeluju 22. Najbogatiji ekosistem raznovrsnošću vrsta na našoj planeti 23. ekološka grupa živih organizama koji stvaraju organske tvari 24. ekološka grupa živih organizama 24. ekološka grupa živih organizama. organizmi koji konzumiraju gotove organske tvari, a ne vrše mineralizaciju 25. ekološka grupa živih organizama koji konzumiraju gotove organske tvari i doprinose njihovoj potpunoj transformaciji u mineralne tvari 26. korisna energija prelazi na sljedeći trofički (nutritivni) nivo 27. potrošači prvog reda 28. potrošači drugog ili trećeg reda 29. mjera osjetljivosti zajednica živih organizama na promjene u određenim uslovima 30. sposobnost zajednica (ekosistema ili biogeocenoza) da održe svoju postojanost i odupru se promjeni životne sredine uslovi 31. niska sposobnost samoregulacije, raznovrsnost vrsta, korišćenje dodatnih izvora energije i visoka produktivnost su karakteristični za 32. veštačku biocenozu sa najvećom stopom metabolizma po jedinici površine. 33. obradive površine zauzimaju 34. gradove zauzima 35. školjka planete naseljena živim organizmima 36. autor doktrina biosfere 37. gornja granica biosfere 38. granica biosfere u dubinama okeana. 39 donja granica biosfere u litosferi. 40. međunarodna nevladina organizacija osnovana 1971. godine, koja sprovodi najefikasnije akcije u odbrani prirode.
A 1. Kriterijum vrste, uzimajući u obzir ukupnost faktora životne sredine u kojima vrsta postoji:Morfološki;
fiziološki;
Geografski;
Ekološki.
A 2. Elementarna struktura na čijoj se razini manifestira djelovanje prirodne selekcije je:
Individualni organizam;
Populacija;
biocenoza;
A 3. Prirodna selekcija za razliku od veštačke:
Pomaže u očuvanju korisnih svojstava za tijelo;
Osigurava očuvanje pojedinaca sa osobinama korisnim za ljude;
Usmjeren na stvaranje i poboljšanje sorti i rasa;
Na snazi je od pojave poljoprivrede i stočarstva.
A 4. Rezultati evolucije uključuju:
Nasljedna varijabilnost;
Borba za postojanje;
fitness;
Prirodna selekcija.
A 5. Primjeri paleontoloških dokaza evolucije su:
Otisci i fosili drevnih oblika života;
Prisutnost rudimentarnih udova kod kitova;
Znakovi gmizavaca u strukturi platipusa;
Znakovi sličnosti u embrionima sisara i riba u ranim fazama razvoja.
A 6. Primjer aromorfoze u crnogoričnim biljkama je pojava u njima:
A 7. Degeneracija uključuje:
Gubitak većine organa od raka sacculina hilar;
Pojava srca sa četiri komore kod ptica;
Pojava krzna kod sisara;
Formiranje ravnog oblika tijela kod raža.
A 8. Kod ljudi, za razliku od majmuna:
Postoje mišići lica;
Postoje prednji udovi sa noktima;
Konstantna tjelesna temperatura;
Kičma ima četiri krivine.
A 9. Primjer biološkog faktora u ljudskoj evoluciji je:
sposobnost za rad;
Upotreba odjeće;
Komunikacija putem usmenog i pismenog govora;
Sposobnost prenošenja stečenih karakteristika nasleđivanjem.
A 10. Vrši se razvoj novih sorti i rasa biljaka i životinja...
Odabir
Genetika
fiziologija
Citologija
pomozi mi molim te??
1. Oplemenjivanje novih sorti biljaka i pasmina životinja vrši se:A – genetika;
B – selekcija;
B – agrobiologija;
G – botanika.
2. Nasljednost je svojstvo organizama:
A – interakcija sa okolinom;
B – odgovoriti na promjene životne sredine;
B – prenijeti svoje osobine i razvojne karakteristike na potomstvo;
D – stiču nove karakteristike u procesu individualnog razvoja.
3. Za proučavanje prirode nasljeđivanja nekoliko osobina od strane nekoliko generacija biljaka i životinja, vrši se ukrštanje:
A – monohibrid;
B – analiziranje;
B – polihibrid;
G – blisko povezan.
4. “Razdvajanje za svaki par karakteristika se dešava nezavisno od drugih parova karakteristika” - ovo je formulacija:
A – Mendelov prvi zakon;
B – Morganov zakon;
G – Mendelov drugi zakon;
D - Mendelov treći zakon.
5. Pojava jedinki sa istim genotipom u prvoj hibridnoj generaciji je manifestacija:
A – zakon cijepanja;
B – pravo samostalnog nasljeđivanja;
B – pravila uniformnosti;
G – zakon vezanog nasljeđivanja.
6. Na sl. prikazuje roditeljske forme u kojima crvena boja lala lala dominira nad bijelom. Kakav će biti genotip potomstva za ovu osobinu ako je roditeljski organizam sa dominantnim osobinama homozigotan?
A – AA;
B – aa;
B – Aaa;
G – Aa.
7. Prema sl. Odredite genotip potomstva (F1) zamorčića ako je poznato da je roditelj s crnom i kratkom dlakom heterozigotan za oba svojstva:
A – AaBv;
B – aavv;
B – aaBv;
G – Aavv.
8. Genotip je ukupnost:
A – spoljašnji znaci tela;
B – unutrašnji znaci tela;
B – geni koje je potomstvo primilo od roditelja;
D – reakcije organizma na uticaje okoline.
9. Srednja priroda nasljeđivanja karakteristika se manifestira kada:
A – postoji promjena uslova okoline;
B – sezonske promjene se dešavaju u prirodi;
B – heterozigoti se po izgledu ne razlikuju od homozigotnih;
D – heterozigoti se po izgledu razlikuju od homozigotnih.
10. Geni koji se nalaze na istom hromozomu:
A – nasleđuje se nezavisno;
B – ulaze u različite zametne ćelije tokom procesa mejoze;
B – naslijeđeno zajedno;
D – dati rascjep u potomstvu u omjeru 3:1.
11. Koje slovo označava fenotip organizma prikazanog na sl.
B – AaBbCc;
B – AbC;
G -
12. Ukrštanje jedinki koje se razlikuju po dva para karakteristika naziva se:
A – polihibrid;
B – analiziranje;
B – dihibrid;
G – monohibrid.
13. Sa genetske tačke gledišta, nasledne bolesti kod ljudi su:
A – izmjene modifikacije;
B – promjena fenotipa nije povezana sa promjenom genotipa;
B – mutacije;
G – odgovor na promene u okruženju, nezavisno od genotipa.
14. Osnova citogenetičke metode proučavanja ljudskog naslijeđa je proučavanje:
A – porodični pedigre;
B – distribucija osobine u velikoj populaciji ljudi;
B – hromozomski set, pojedinačni hromozomi;
D – razvoj simptoma kod blizanaca.
15. Promjena redoslijeda nukleotida u molekuli DNK naziva se:
A – mutacije gena;
B – hromozomske mutacije;
B – somatske mutacije;
G – kombinativna varijabilnost.
16. Granice unutar kojih su moguće modifikacije određene karakteristike nazivaju se:
A – prilagodljivost;
B – norma reakcije;
B – varijabilnost;
G – razdražljivost.
17. Pod uticajem genotipa i uslova sredine formira se:
A – norma reakcije;
B – nasljednost;
B – fenotip;
G – fitnes.
18. Izolacija iz izvornog materijala čitave grupe jedinki sa karakteristikama neophodnim za uzgajivača naziva se:
A – prirodna selekcija;
B – masovna selekcija;
B – individualni oblik vještačke selekcije;
G – spontana selekcija.
Čini se da je definisanje pojma „vrste“ veoma teško.
Vrstu ćemo smatrati istorijski uspostavljenim skupom organizama koji zauzimaju određeno stanište i koje karakterizira zajedničko porijeklo, sličan sistem prilagođavanja na nivoe životne sredine i reprodukcija osnovnih adaptivnih osobina i karakteristika tokom generacija.
Organizmi jedne vrste imaju fenotip i genotip karakteristične za tu vrstu koji se razlikuju od organizama druge vrste.
Vrsta zauzima određeni raspon. Područje distribucije nekih vrsta je široko, a takve vrste su obično politipne - uključuju nekoliko geografskih rasa ili podvrsta. Druge vrste imaju mnogo ograničeniji raspon, po pravilu ne formiraju geografske rase i monotipne su. Jedinke koje čine vrstu ne čine stalnu, homogenu masu. Svaki organizam vrste, koji ima opća i karakteristična svojstva, ima i svoje individualne genotipske karakteristike, koje zajedno predstavljaju nasljednu varijabilnost vrste, ili, kako se ponekad naziva, „genofond“ vrste.
Vrsta, koja predstavlja jedan skup organizama, podijeljena je u zasebne populacije. Populacija naziva se skup jedinki iste vrste koje se slobodno ukrštaju, koje karakterizira zajedničko stanište i prilagođavanje datim uvjetima postojanja. Populacija se formira pod uticajem uslova postojanja zasnovanih na interakciji faktora naslednosti, varijabilnosti i selekcije. Formiranje populacija je jedinstven način „prilagođavanja“ vrste specifičnim uslovima njenog postojanja. Životinjske pasmine i biljne sorte stvorene umjetnom selekcijom također su zastupljene zasebnim populacijama.
Procesi formiranja populacije i njihova dinamika čine mikroevoluciju. Pojava novih vrsta počinje divergencijom – podjelom vrste na zasebne, neukrštajuće ili izolirane grupe organizama. Populacija je neka vrsta „kovanice“ u kojoj prirodna selekcija stvara nove oblike.
U prirodi, populacije svake vrste karakterizira genetska raznolikost. Ali to često ne primjećujemo. Pojedinci populacije i vrste izgledaju nam relativno ujednačeni izgledom. Ova relativna uniformnost životinja i biljaka, koja omogućava taksonomistima da klasifikuju životinje i biljke kao određene vrste, podvrste i rase, stvorena je prirodnom selekcijom. Selekcija osigurava ne samo raznolikost, već i uniformnost unutar vrste.
Međutim, ova uniformnost se odnosi samo na glavne tipične karakteristike, karakteristike i svojstva organizama u datoj populaciji. Čim počnemo genetski analizirati sastav populacije do detalja, rastavljajući je u zasebne linije, odmah ćemo otkriti ogromnu genotipsku varijabilnost. Ispada da svaka populacija, unutar koje su se jedinke ukrštale dugo vremena, na ograničenom području pod datim klimatskim uslovima, ima svoj karakter varijabilnosti.
Izvori nasljedne varijabilnosti u populaciji su mutacijska i kombinacijska varijabilnost. Djelovanje genetskih zakona u populaciji je predmet proučavanja populacione genetike.
U prirodi ne postoje dva organizma koja su apsolutno slična po genotipu. Zoolozi ili botaničari koji proučavaju biološke procese u bilo kojoj grupi jedinki uvijek imaju posla s nasljedno heterogenom grupom organizama. Ali budući da proučavaju fenotip organizama, imaju pravo zanemariti genetsku raznolikost svog materijala.
Proučavanje populacije može se provesti čisto deskriptivnom metodom. U ovom slučaju utvrđuju se fenotipske karakteristike oblika populacije, njene biološke karakteristike, razjašnjavaju se uslovi postojanja i odnosi između organizama, lanci ishrane, konkurencija, dinamika populacije kroz godine i njena zavisnost od različitih faktora. Populacije su izolirane i formirane kao rezultat djelovanja mnogih faktora: načina razmnožavanja, prirode varijabilnosti, promjena u broju jedinki, tempa i smjera selekcije, klimatske, geografske i fiziološke izolacije. Glavni je odabir osobina koje osiguravaju proces reprodukcije generacija, odnosno reprodukciju. Očigledno je da se kod različitih metoda razmnožavanja nastanak i održavanje populacija odvija na različite načine, što se može vidjeti poređenjem populacija unakrsnih (alogamnih) i samooplodnih (autogamnih) organizama.
Za postojanje populacije od najveće su važnosti različite vrste nasljedne varijabilnosti: mutacije gena, hromozomski preustroj i poliploidija. Nenasljedne promjene mogu igrati samo ograničenu ulogu. Organizmi koji se genotipski razlikuju, na primjer, po jednom genu, možda se ne razlikuju morfološki jedni od drugih, ali imaju različite fiziološke karakteristike (živost, trajanje razvoja, plodnost). Genetske metode omogućavaju da se dobije potpunija slika o nasljednom potencijalu populacije, njenim adaptivnim karakteristikama i smjeru evolucije.
Pioniri proučavanja genetike moraju biti prepoznati kao oplemenjivači, jer je za identifikaciju raznolikosti genotipova u populacijama bilo potrebno izolovati i odabrati za ukrštanje pojedine roditeljske parove i potom proučavati njihovo potomstvo u nizu generacija. Upravo to su radili uzgajivači kada su stvarali razne sorte i rase. Međutim, naučni temelji genetskog proučavanja populacije mogli su biti postavljeni tek nakon otkrića G. Mendela, koji je ustanovio kvantitativne obrasce nasljeđivanja.
Prvo naučno istraživanje populacije, kombinujući genetičke i statističke metode, preduzeo je danski biljni fiziolog i genetičar V. Johannsen. Njegovo klasično djelo O nasljeđivanju u populacijama i čistim linijama, objavljeno 1903., označilo je početak genetskog proučavanja populacija. Kao što se često dešava u nauci, klasično otkriće dolazi na naizgled elementarnom fenomenu i korišćenjem jednostavne tehnike. Tako je u ovom slučaju V. Johannsen za predmet proučavanja odabrao populacije ne unakrsno oprašujućih, već samooprašujućih biljaka - ječam, pasulj i grašak. Metodološki, ovo je pojednostavilo rad, jer se svaka takva populacija lako mogla podijeliti u grupe potomaka pojedinih jedinki, odnosno identificirati zasebne, „čiste“ linije. „Čistom linijom nazivam“, napisao je, „pojedince koji potiču od jedne jedinke koja se samooprašuje. Iz ovoga je jasno da se populacija apsolutnih samooprašivača sastoji samo od čistih linija, čije se jedinke u prirodi mogu miješati, ali na njih ne može utjecati ukrštanje.”
Težina i veličina sjemena su uzete kao karakteristike. Ove kvantitativne osobine određene su djelovanjem mnogih gena, odnosno određene su poligenski, te su vrlo podložne varijabilnosti pod utjecajem okolišnih faktora - sastav tla, klima, način sadnje itd. Stoga treba utvrditi prirodu njihovog nasljeđivanja. , potrebno je koristiti matematičke metode analize varijabilnosti.
Za ove karakteristike postoji izražena modifikacija, ili paratipska, varijabilnost. Postojala su različita gledišta o značaju ove varijabilnosti za evoluciju u biologiji. Zagovornici teorije nasljeđivanja stečenih svojstava vjerovali su da se promjene uzrokovane uticajem faktora sredine nasljeđuju i prenose na potomstvo. Protivnici ove teorije poricali su nasljeđivanje modifikacijskih promjena. Rješenje ovog spora u korist potonjeg bilo je od fundamentalne važnosti, jer je selekcija organizama po fenotipu bez utvrđivanja nasljednih potencijala ranije bila široko rasprostranjena u selekciji i kočila je razvoj životinjskih pasmina i biljnih sorti.
Johannsen je izmjerio sjeme jedne sorte pasulja i napravio niz varijacija za ovaj indikator. Utvrđeno je da je težina sjemena promjenjiva u rasponu od 150 do 750 mg. Zatim je sjeme mase od 250 do 350 mg posijano odvojeno od sjemena mase 550 do 650 mg. Sjeme svake uzgojene biljke ponovo je izvagano. Budući da je pasulj samooprašujuća biljka, genotip sjemena jedne biljke trebao bi biti isti, ali sjemenke različitih biljaka mogu imati genotipske razlike. Stoga je teško sjeme (550-650 mg) i lagano sjeme (250-350 mg) odabrano od sorte reprezentativne za populaciju dalo biljke čije je sjeme značajno variralo u težini. Prosječna težina sjemena na biljkama uzgojenim iz teškog sjemena iznosila je 518,7 mg, a na biljkama uzgojenim iz lakog sjemena 443,4 mg. Ovo je pokazalo da se sorta pasulja-populacija sastoji od genetski različitih biljaka, od kojih svaka može postati predak čiste linije.
Tokom 6-7 generacija, Johannsen je selekcionirao i teško i lagano sjeme iz svake biljke posebno, odnosno birao je unutar čistih linija. Takvom selekcijom nije došlo do pomaka u nizu generacija prema teškim ili lakim sjemenkama ni u jednoj liniji. Posljedično, varijabilnost u težini sjemena unutar čiste linije bila je nenasljedna i modificirana.
Kao rezultat svog istraživanja, Johannsen je došao do sljedećih zaključaka: 1) “selekcija u populaciji uzrokuje ... veći ili manji pomak - u smjeru selekcije - prosječne osobine oko koje, fluktuirajući, variraju odgovarajuće jedinke ” i 2) „unutar čistih linija regresija (stepen sličnosti osobine potomstva sa majčinom) bila je... potpuna; selekcija unutar čistih linija nije izazvala nikakvu promjenu tipa.”
Kao što vidimo, populaciju autogamnih biljaka čine genotipski heterogene linije. Biljke takve populacije se ne križaju i ne razmjenjuju nasljedne informacije. U ovom slučaju, postojanje populacije zasniva se na strogoj prirodnoj selekciji linija određenog genotipa, na zajedništvu mehanizama prilagođavanja sličnim uslovima sredine. Drugim riječima, promjene u populaciji autogamnih biljaka i životinja provode se selekcijom određenih nasljedno različitih linija i klonova koji imaju adaptivne prednosti.
Prilikom samooplodnje pojedini organizam može biti osnivač nove rase, podvrste i vrste, kao i sorte ili rase. Na primjer, nova sorta pšenice može se razviti iz jednog zrna odabranog iz populacije.
Međutim, govoreći o visokoj homozigotnosti čistih linija, treba imati na umu da ni čiste linije ne mogu biti apsolutno homozigotne iz sljedećih razloga. Prvo, ne postoje obavezne (apsolutne) samooplodne biljke. U populacijama samooprašivača, na primjer pšenice, paradajza itd., biljke s otvorenim cvjetanjem i unakrsnom oprašivanjem uvijek se nalaze s različitom učestalošću. Zbog toga se u populacijama, iako rijetko, dešavaju procesi ukrštanja i, shodno tome, razmjene nasljednih informacija između čistih linija. Drugo, biljke koje se samooprašuju imaju mutacije koje sprečavaju samooprašivanje (nekompatibilnost). Treće, u čistim linijama samooprašivača, čak i u jednoj generaciji, javlja se vrlo primjetan broj raznolikih mutacija koje narušavaju homogenost čiste linije.
Zbog ovih razloga, sorte samooprašujućih biljaka, kada se razmnožavaju u proizvodnji, mogu izgubiti neke od svojih sortnih kvaliteta i zahtijevaju stalno praćenje, što je osnova za potrebu obnavljanja sorte.
Prilikom vegetativnog razmnožavanja agamskih organizama koji nemaju polni proces ili su ga drugi put izgubili (neke protozoe, gljive, alge itd.), pojedinačni klonovi su predmet selekcije u populaciji. Genetski integritet (integracija) takvih klonova u populaciju je vrlo nizak zbog nemogućnosti ukrštanja jedinki različitih klonova, ali takve populacije očigledno još uvijek postoje u prirodi i održavaju se selekcijom na temelju simbiotskih odnosa različitih genotipova.
Kod unakrsnih oplodnih organizama u prirodi populacija se formira na osnovu slobodnog ukrštanja jedinki suprotnog pola različitih genotipova, odnosno na osnovu panmiksije. U ovom slučaju, nasljedna struktura sljedeće generacije se reprodukuje na osnovu različitih kombinacija različitih gameta tokom oplodnje. Iz toga slijedi da će broj jedinki određenog genotipa u svakoj generaciji biti određen učestalošću pojavljivanja različitih gameta koje proizvode genotipski različiti roditeljski organizmi. To znači da se osobine i svojstva čuvaju i distribuiraju u populaciji na osnovu obrazaca promjena u učestalosti distribucije gena. Ove promjene su zasnovane na obrascima nasljeđivanja koje su otkrili G. Mendel i T. Morgan. Poznavanje ovoga omogućilo je izvođenje pravila za distribuciju gena u panmiktičkoj populaciji.
Očigledno je da će organizmi čiji genotipovi najbolje osiguravaju adaptaciju na uvjete postojanja proizvesti veći broj odgovarajućih gameta od onih manje prilagođenih. Shodno tome, učestalost (pojava) određenog gena u populaciji će također biti određena prirodnom selekcijom.
Neki genetičari zajednicu slobodno ukrštanja genotipski različitih organizama unutar vrste nazivaju mendelovskom populacijom. Radije ga nazivamo panmiktičkim, jer je njegovo postojanje određeno ne samo Mendelovim zakonima, već i interakcijom svih evolucijskih faktora koji osiguravaju slobodu ukrštanja organizama unutar populacije. Raznolikost genotipova u panmiktičkoj populaciji rezultat je mutacijske i kombinacijske varijabilnosti. Novonastala mutacija, da bi postala vlasništvo populacije, mora opstati i umnožiti se, odnosno postati dio genotipova brojnih organizama. Svaka mutacija u populaciji ima svoju sudbinu.
Zbog širenja velikog broja različitih mutacija u populaciji, genotipovi organizama postaju zasićeni raznim mutacijama, koje su najčešće u heterozigotnom stanju. Na primjer, broj biljaka heterozigotnih za određene mutacije može biti prilično visok postotak u populaciji. Kako se koncentracija mutacija u populaciji povećava, one postaju homozigotne.

Ogromna zasićenost populacije mutantnim genima karakteristična je ne samo za kultivisane biljke i domaće životinje, već, kako je prvi pokazao S.S. Chetverikov, i za prirodne populacije. Istovremeno, u populaciji se javljaju mutacije koje se razlikuju kako po svojoj genetskoj prirodi (genske mutacije i kromosomski preuređenje) tako i po fenotipskoj manifestaciji.
Da bismo ilustrirali podjelu panmiktičke populacije pod utjecajem selekcije, razmotrimo model eksperimenta s umjetno stvorenom hibridnom populacijom, koji su izveli američki genetičari D. Jones i E. East. Ova dva istraživača su ukrstila dvije vrste duhana koje su se razlikovale po dužini vjenčića (kratak i dug). Biljke prve generacije su ukrštane jedna s drugom, a iz druge generacije su uzete dvije linije A i B sa sličnom varijabilnosti po ovom svojstvu.
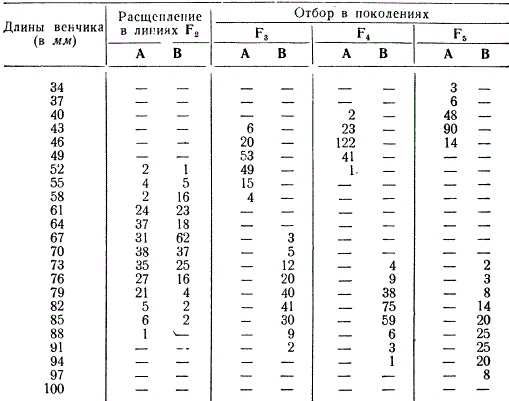
Duljinu vjenčića određuju mnogi geni, pa se u F 2 kretala u rasponu od 52 do 88 mm u ovim linijama. Naknadno je izvršena selekcija u potomstvu uzetih linija u tri generacije: u liniji A - na kratko. vjenčić, a u liniji B - na dugoj metlici. U svakoj generaciji odabrani oblici su ukrštani unutar obje linije: u liniji A - sa kratkim vjenčićem, au liniji B - sa dugim vjenčićem. Kao što vidimo, već u petoj generaciji linije A i B bile su toliko različite da između njih nije bilo prijelaza (transgresije), tj. maksimalna dužina vjenčića u liniji A bila je manja od minimalne dužine u liniji B.
Shodno tome, selekcijom i ukrštanjem odabranih oblika moguće je stvoriti linije sa drugačijim izrazom osobine od one u izvornoj populaciji: selekcija dijeli populaciju na različite genotipove. U ovom eksperimentu izvršena je umjetna selekcija za jedno svojstvo uz namjerno ukrštanje biljaka. U prirodi se prirodna selekcija provodi prema mnogim karakteristikama i ili čuva i održava populaciju u integralnom stanju, ili je razlaže prema specifičnim uslovima postojanja.
Proučavanje populacijske genetike provodi se različitim metodama, a glavne su citogenetske, ekološko-fiziološke i matematičke.
Prve dvije metode se koriste u analizi nasljeđivanja u populaciji - za procjenu koncentracije mutacija i učestalosti mutacija. Pokazalo se da je ekološko-fiziološka metoda neophodna za procjenu djelovanja abiotičkih i biotičkih faktora na određivanje adaptivne vrijednosti fenotipova koji pripadaju genetski različitim klasama jedinki. Istovremeno, otvaraju se velike eksperimentalne mogućnosti modeliranjem djelovanja selekcije u umjetno stvorenim sintetičkim populacijama sa unaprijed određenim genetskim parametrima - unošenje određenih mutacija, inverzija, translokacija itd. u populaciju.
Matematički metod nam omogućava da damo strog kvantitativni opis bioloških procesa. Upotreba elektronskih računara pokazala se posebno obećavajućom za modeliranje dinamike genetske strukture populacije, uzimajući u obzir složenu interakciju mnogih faktora. Situacije opisane u ovom slučaju znatno su bliže odražavanju prave, složene i kontradiktorne slike evolucijskih procesa koji se dešavaju u prirodnim populacijama biljaka i životinja.
Ako pronađete grešku, označite dio teksta i kliknite Ctrl+Enter.
Vježba 1. Popunite prazna polja u datim izjavama.
1. Vještačka populacija jedinki jedne vrste, koju je čovjek uzgojio za određenu praktičnu svrhu, naziva se... ili....
2. Stvorena je zbirka svjetskih sorti pšenice... i pohranjena u... .
3. Najtradicionalnije metode selekcije su... i...
4. Najefikasniji oblik selekcije je... selekcija.
5. Prisilno samooprašivanje biljaka koje se međusobno oprašuju naziva se....
6. Inbreeding je... uzgoj domaćih životinja.
7. Najčešći vid ukrštanja u uzgoju životinja je....
8. Hibridi iz ukrštanja genetski različitih roditeljskih oblika odlikuju se... .
9. Udaljeni hibridi i kod biljaka i kod životinja, po pravilu, ... .
10. Tritikale je alopoliploidni hibrid pšenice sa... .
11. Somatska hibridizacija je hibridizacija na... nivou.
12. Transgene su osobe čiji genom sadrži... genetske informacije.
13. Genetski homogeno potomstvo dobijeno od jedne jedinke naziva se....
Kontrolna pitanja
1. U koje svrhe se samonikle biljne vrste koriste kao izvorni materijal za selekciju?
2. Šta je uvod?
3. Za koje osobine uzgajivač razvija nove sorte i rase?
4. Poznavanje kog zakona pomaže uzgajivaču u potrazi za spontanim mutacijama?
5. Šta je bio podsticaj za razvoj selekcije zračenja?
6. U koju svrhu se zrače sjeme, mjesta rasta i drugi dijelovi biljaka?
7. Koje vrste varijabilnosti koristi uzgajivač pri stvaranju novih oblika biljaka i životinja?
8. Šta je incest?
9. U koju svrhu se stvaraju inbred linije?
10. Šta je heteroza i koji je njen mehanizam?
11. Šta je prepreka udaljenoj hibridizaciji?
12. Koji su načini da se prevaziđe neukrštanje biljaka?
13. Šta je uzrok neplodnosti kod udaljenih hibrida?
14. Kako se masovna selekcija razlikuje od individualne selekcije?
15. Koja je selektivna prednost poliploidnih biljnih oblika?
16. Koje biljke se nazivaju amfidiploidi?
17. Koji je mehanizam djelovanja kolhicina?
18. Koja je svrha dobijanja haploidnih biljaka?
19. Šta se podrazumijeva pod pojmom „zelena revolucija“?
20. Kako se proizvode transgene biljke?
21. Šta je somatska hibridizacija i u koju svrhu se izvodi?
22. Kakvi su izgledi za kloniranje životinja?
23. Koje domaće uzgajivače poznajete?
Prilikom pripreme za ispit iz biologije bit će od koristi i sljedeći materijali iz knjige:
ALLEL - jedno od stanja istog gena. Postoje dominantni (A) i recesivni (a) aleli.
ALLELIC GENES– geni koji određuju alternativni razvoj iste osobine i nalaze se u identičnim regijama homolognih hromozoma.
ALTERNATIVNI ZNAKOVI- znakovi koji se odnose na istu imovinu, ali različito izraženi.
AUTOSOMI- hromozomi koji su isti kod muškaraca i žena
BIVALENT- par konjugiranih homolognih hromozoma.
GENE- dio nukleinske kiseline koji je jedinica funkcije i može se promijeniti mutacijom.
GENETIKA- nauka o zakonima naslijeđa i varijabilnosti organizama.
GENOME- skup gena sadržanih u haploidnom setu hromozoma.
HETEROSIGOTNI INDIVIDUALCI- jedinke sa različitim alelima istog gena (Aa).
AaBv - genotip diheterozigotne jedinke; AaBbCs - genotip triheterozigotne jedinke
HETEROCHROMOSOMES(ili spolni hromozomi) - hromozomi po kojima se muški i ženski organizmi razlikuju jedni od drugih.
(XX – homogametno; XY – heterogametno)
HOMOZIGOTNE INDIVIDUALNOSTI- jedinke sa istim alelima istog gena.
Postoje dvije vrste homozigota: dominantni, kada su oba alela gena dominantna (AA) i recesivna, kada su oba alela recesivna (aa).
HOMOLOGIJA HROMOSOMI- hromozomi koji su identični po veličini i morfologiji, koji sadrže isti broj gena odgovornih za iste karakteristike i svojstva organizma.
Unatoč svim sličnostima homolognih hromozoma, oni su, u pravilu, genetski neidentični, jer geni koji se nalaze u njima, a koji određuju iste karakteristike, mogu biti predstavljeni različitim alelima. Jedan homologni hromozom može imati dominantni alel gena (A), a drugi može imati recesivni alel istog gena (a).
DIHYBRIDNO KRŠTANJE- ukrštanje roditeljskih jedinki koje se razlikuju po dvije karakteristike
DOMINANTNI ALEL GENA- alel čije djelovanje potiskuje djelovanje drugog recesivnog alela istog gena.
DOMINANTNI KARAKTER - osobina čiji razvoj kod hibridnih jedinki potiskuje razvoj druge recesivne osobine.
VARIJABILNOST
KODOMINACIJA- manifestacija oba alela u fenotipu heterozigotnih jedinki (Aa).
CROSSINGOVER- razmjena sekcija između homolognih hromozoma tokom njihovog
konjugacija.
MONOHIBRIDNO UKRŠĆANJE– ukrštanje roditeljskih jedinki koje se razlikuju po jednoj osobini
NASLJEDNOST- svojstvo organizama da osigurava materijalni i funkcionalni kontinuitet između generacija.
POLIMERIZAM- takva interakcija nealelnih gena u kojoj je razvoj osobine određen interakcijom dva ili više parova gena koji imaju sličan učinak na razvoj ove osobine.
RECESIVNI ALEL GENA
VEZANO NASLJEĐE(MORGANOV zakon) - geni lokalizovani na jednom hromozomu su povezani i prenose se pretežno zajedno.
KVAČILNA GRUPA– geni lokalizovani na istom hromozomu (zajedno naslijeđeni)
FENOTIP- skup spoljašnjih i unutrašnjih karakteristika i svojstava organizma, determinisanih interakcijom genotipa sa određenim uslovima sredine.
HROMATIDI– jedan od dva nukleoproteinska lanca formirana tokom udvostručavanja hromozoma tokom ćelijske deobe (ili ćerki hromozoma)
HROMOSOMI- samoreproducirajuće organele ćelijskog jezgra, koje su nosioci gena koji određuju nasljedna svojstva ćelija i organizama.
CENTROMERE- dio hromozoma koji drži svoja 2 lanca zajedno (hromatide)
EPISTASE- ovo je interakcija između nealelnih gena, u kojoj je djelovanje jednog gena potisnuto drugim
Amitoza- direktna dioba stanica, kada se interfazno jezgro dijeli ligacijom bez stvaranja struktura karakterističnih za kariokinezu.
OUTBRIDING- ukrštanje pojedinaca koji nisu u bliskom srodstvu. Suprotnost izvanbredingu je inbriding.
BLASTOMERES- ćelije nastale kao rezultat fragmentacije zigota na samom početku razvoja embrija.
POGLED- istorijski uspostavljen skup jedinki, ujedinjenih mogućnošću međusobnog ukrštanja, koji zauzimaju određeno stanište, koje karakteriše zajedničko poreklo i sličan sistem prilagođavanja uslovima životne sredine.
Vrsta je genetski zatvoren sistem, budući da se jedinke različitih vrsta normalno ne križaju, stoga nema razmjene genetskih informacija.
HAPLODIJA - vidi genomske mutacije.
GENEALOŠKA METODA- metoda zasnovana na sastavljanju i analizi rodovnika koji ukazuju na ispoljavanje karakteristika od interesa za istraživača. Ova metoda se koristi u genetskom savjetovanju, kao iu uzgoju životinja. Analizom pedigrea moguće je utvrditi prirodu nasljeđivanja osobina u mnogim slučajevima, moguće je utvrditi latentno nosivost gena za bilo koje nasljedne bolesti, utvrditi genotipove osoba navedenih u rodovniku i odrediti postotak; vjerovatnoća da će u porodici sa nasljednim opterećenjem imati bolesno dijete.
GENOMSKIH MUTACIJA- mutacije koje nastaju kao rezultat promjena u broju hromozoma u kariotipu pojedinca.
Višestrukom haploidnom promjenom broja hromozoma nastaju poliploidne ili haploidne jedinke. Kada promjena u broju hromozoma nije višestruka od haploidnog, nastaju aneuploidni organizmi. Na primjer, kod Downovog sindroma ljudi nemaju 46 hromozoma, već 47, dodatni hromozom u 21. paru, odnosno ne postoje dva, već tri hromozoma. Drugim riječima, trisomija na hromozomu 21 uzrokuje ovu nasljednu bolest.
GENOTIP- sistem gena koji su u interakciji lokalizovani u hromozomima jezgra somatskih ćelija.
GENE POOL- kompletan skup gena karakterističnih za određenu vrstu, formiran u procesu njene evolucije.
Termin genofond se koristi ne samo u odnosu na vrstu, već iu odnosu na populaciju.
Genofond jedne populacije treba shvatiti kao kompletan skup gena karakterističnih za određenu populaciju organizama.
HERMAFRODIT- organizam koji ima razvijen i funkcionalan muški i ženski reproduktivni sistem, formirajući muške i ženske gamete.
Najčešće se pravi hermafroditizam javlja kod cvjetnica (sve jednodomne biljne vrste su hermafroditi), rjeđe kod životinja (hidra, glista itd.).
HETEROZIS- (hibridna snaga) svojstvo hibridnih organizama da nadmaše svoje roditelje u produktivnosti, vitalnosti i veličini tijela.
Fenomen heterozisa se koristi u poljoprivrednoj praksi korišćenjem semena hibridnih biljaka za setvu, što rezultira povećanjem prinosa. Za setvu useva kao što je kukuruz koristi se gotovo samo hibridno seme.
IDIGRAM HROMOSOMA - grafički prikaz hromozoma kariotipa, raspoređenih u određenom redoslijedu od velikog prema malom ili obrnuto.
Embryo Sac- ženski gametofit kod kritosjemenjača.
ZYGOTE- oplođeno jaje koje daje povod za razvoj novog organizma.
DOWN SINDROM- nasljedna bolest uzrokovana prisustvom viška hromozoma (trizomija 21. hromozoma) ukupno umjesto 46 kod bolesnih osoba - 47 hromozoma. Bolesnike karakterišu sljedeća odstupanja: ukošene oči, veliki debeo jezik, nisko postavljene, deformirane i naprijed okrenute uši, mentalna retardacija, niska inteligencija itd. Stopa nataliteta djece sa Downovim sindromom je 1:600.
DELECTION- hromozomska mutacija koja nastaje kao rezultat lomova hromozoma i gubitka dijela nasljednog materijala.
Postoje intrahromozomske delecije, koje nastaju dvostrukim prekidom na tijelu hromozoma i gubitkom izrezane dionice, te terminalni nedostaci (deficijencije).
DRIFT OF GENEES- promjena frekvencije gena u populaciji uslijed djelovanja nasumičnih faktora.
Genetski drift (ili genetski-automatski procesi) karakterističan je za male populacije organizama.
Uzroci genetskog drifta mogu biti prirodne katastrofe: poplave, šumski požari, epizootije, masovno širenje štetočina ili utjecaj ljudskih aktivnosti (krčenje šuma, velika upotreba pesticida, itd.). Naglo smanjenje broja jedinki u populaciji, pored promjena u frekvenciji gena, dovodi do povećanja inbreedinga, povećane homozigotnosti jedinki i smanjenja nasljedne varijabilnosti u populaciji.
ZAKON VAVILOVA N. I. - (zakon homoloških nizova u nasljednoj varijabilnosti) - genetski bliske vrste i rodove karakteriziraju slični nizovi nasljedne varijabilnosti sa takvom pravilnošću da se, poznavajući broj oblika unutar jedne vrste, može pretpostaviti prisustvo paralelnih oblika unutar druge vrste i rodovi.
Zakon N.I. Vavilova promiče veći fokus u radu uzgajivača i u skladu je sa učenjem Charlesa Darwina o paralelnoj varijabilnosti.
Na primjer, crna boja zrna nalazi se u raži, pšenici, ječmu, kukuruzu i drugim žitaricama; izduženi oblik zrna nalazi se u svim žitaricama.
HARDY-WEINBERGOV ZAKON- relativne frekvencije gena u populaciji se ne mijenjaju iz generacije u generaciju u uvjetima idealne populacije
Posljedica Hardy-Weinbergovog zakona je očuvanje omjera homo- i heterozigota u populacijama.
Hardy-Weinbergov zakon je karakterističan za velike populacije i određuje genotipsku ravnotežu u tim populacijama. U prirodnim uslovima, jasna manifestacija Hardy-Weinbergovog zakona može biti narušena kao rezultat procesa mutacije i akcije selekcije. Stoga, u prirodnim populacijama postoji samo tendencija da se održi odnos genskih alela i genotipske ravnoteže.
VARIJABILNOST- svojstvo organizama da steknu nove karakteristike koje ih razlikuju od roditelja i jedni od drugih.
Dakle, varijabilnost uzrokuje nestabilnost u očuvanju osobina i svojstava roditelja od strane potomstva. Postoji genotipska i fenotipska varijabilnost. GENOTIPIČNO- varijabilnost zbog promjena u strukturi genotipa.
Genotip uključuje mutacijsku i kombinativnu varijabilnost (vidi mutacija, kombinativna varijabilnost).
KOMBINATIVNA VARIJABILNOST- varijabilnost organizama koja nastaje kao rezultat novih kombinacija genskih alela u genotipu jedinki potomaka dobijenih od istog para roditelja tokom seksualne reprodukcije.
Kombinativna varijabilnost je uzrokovana rekombinacijom nasljednog materijala u mejozi, tokom formiranja gameta. Kao rezultat kombinovane varijabilnosti, braća i sestre se međusobno razlikuju po skupu karakteristika i svojstava, a razlikuju se i od svojih roditelja.
VARIJABILNOST IZMJENA- to je varijabilnost u kojoj se pod uticajem uslova sredine fenotip jedinke menja bez promene njenog genotipa.
Varijabilnost modifikacije pokazuje raznolikost u ispoljavanju istih genotipova u različitim uslovima sredine. MI je genotipski određen, tj. granice raspona varijabilnosti osobine su genotipski ograničene (vidi normu reakcije).
ONTOGENETSKA VARIJABILNOST - uzastopne promene kod pojedinca u procesu njegovog rasta i razvoja. Ontogenetska varijabilnost takođe uključuje promene u ćelijama tokom procesa diferencijacije.
FENOTIPSKA VARIJABILNOST- promjena fenotipa jedinke bez promjena u strukturi njenog genotipa. Fenotip uključuje ontogenetsku i modifikacionu varijabilnost (vidi ontogenetsku i modifikacionu varijabilnost).
INBRIDNA DEPRESIJA - smanjenje održivosti i produktivnosti jedinki kao rezultat inbreedinga.
INBRIDING- inbriding jedinki.
Tokom inbreedinga, homozigotnost se javlja kod jedinki potomaka za brojne osobine.
INVERZIJA- intrahromozomska mutacija koja se javlja kao rezultat dva ili više prekida, rotacije hromozomskog preseka za 180°.
Kao rezultat inverzije dolazi do djelomične promjene u redoslijedu gena na hromozomu.
INDIVIDUALNI IZBOR- odabir jedinki procijenjen po potomstvu u nizu generacija.
Tako se prilikom individualne selekcije analizira genotip jedinki. Ova selekcija se koristi pri uzgoju novih rasa životinja i biljnih sorti.
KLON- potomstvo jedne izvorne jedinke, dobiveno njenom aseksualnom reprodukcijom, na primjer, klon ribizle - biljke dobivene ukorjenjivanjem reznica uzetih iz istog grma ribizle.
KLEINFELTEROV SINDROM- nasljedna bolest uzrokovana prisustvom dodatnog spolnog X hromozoma u kariotipu. U kariotipu pacijenata ima 47 hromozoma
VARIJABILNOST
(44 autosoma + XXV). Rod se formira kao muški sa manifestacijom genkomastije, odnosno formira se niz sekundarnih polnih karakteristika prema ženskom tipu - razvoj mliječnih žlijezda, taloženje sala u karlici i butinama, muško primarno i sekundarno polne karakteristike su nedovoljno razvijene. Pacijenti su visoki, imaju smanjene mentalne sposobnosti, povećanu sugestibilnost i lako su pod utjecajem drugih.
KODIRANJE- manifestacija oba alela kod heterozigotnih jedinki (Aa).
Na primjer, četvrta krvna grupa kod ljudi sa AB genotipom. KOMPLEMENTARNO- naziva se interakcija nealelnih gena, čijim se zajedničkim djelovanjem razvija nova osobina koja je odsutna kod roditelja.
KONJUGACIJA- 1) parno privremeno spajanje homolognih hromozoma u mejozi (vidi bivalentno). 2) oblik polnog procesa kod nekih organizama, u kojem dolazi do djelomične razmjene nasljednog materijala (npr. kod dijatomeja, nižih gljiva, cilijata, bakterija).
KOPULACIJA- 1) prodor muške gamete u žensku, spajanje gameta. 2) polni odnos kod životinja sa kopulacionim organima.
KRIS - KRIŽNO NASLJEĐE(unakrsno nasljeđivanje) - naslijeđe vezano za spol, kada se majčina osobina javlja samo kod sinova, a očeva samo kod kćeri.
Unakrsno nasljeđivanje se opaža kod nasljeđivanja vezanog za X, kada majka ima recesivni alel gena na oba X hromozoma, i dominantni alel istog gena na jednom očevom X hromozomu.
CROSSINGOVER- razmjena sekcija tokom konjugacije homolognih hromozoma.
MASOVNI IZBOR- selekcija jedinki po fenotipu bez analize genotipa.
Masovna selekcija se po pravilu koristi za vraćanje ekonomski vrijednih karakteristika sorte, odnosno za osiguranje stabilnosti sortnih karakteristika biljaka.
VIŠESTRUKI ALELIZAM- niz alela istog gena.
Niz alela gena rezultat je ponovljenih mutacija istog gena.
MEJOZA- dioba stanica kako bi se formirale haploidne, genetski neidentične ćelije.
U genetskom smislu, mejoza je mehanizam za distribuciju nasljednog materijala tokom seksualnog razmnožavanja. Genetska suština mejoze je da je nasljedni materijal nejednako raspoređen u kvaliteti i prepolovljen u količini. Posljedica rekombinacije nasljednog materijala u mejozi je neidentičnost nastalih gameta, neidentitet potomaka jedinki dobijenih od istog para roditelja.
MENDELOVI ZAKONI:
1. ZAKON UNIFORMANOSTI (dominacije).
Kod hibrida prve generacije, od par alternativnih osobina, razvija se samo jedna osobina, druga osobina se ne pojavljuje.
2. ZAKON CIJEPANJA.
U drugoj generaciji se uočava cijepanje u određenom kvantitativnom omjeru jedinki s dominantnim i recesivnim osobinama.
Tako, na primjer, monohibridno ukrštanje karakterizira cijepanje u omjeru 3:1, odnosno tri dijela jedinki s dominantnom osobinom i 1 dio s recesivnim. Za dihibrid, formula cijepanja je 9:3:3:1, za trihibrid je 27:9:9:9:3:3:3:1, itd.
3. ZAKON NEZAVISNOG KOMBINACIJE ili samostalnog nasljeđivanja.
Parovi alternativnih osobina se nasljeđuju nezavisno jedan od drugog.
Tako se u poznatom dihibridnom ukrštanju graška u F 2 dobije podjela u omjeru: 9 dijelova žutih okruglih sjemenki 3 dijela žutih naboranih sjemenki 3 dijela zelenih okruglih sjemenki 1 dio zelenih naboranih sjemenki
Ako, analizirajući ove rezultate, pratimo razdvajanje zasebno za svaki par alternativnih karakteristika, nalazimo da u pogledu boje imamo omjer od 12 dijelova žutog prema 4 dijela zelenog sjemena, smanjujući za 4, dobivamo cijepanje od 3: 1. Takođe, bez obzira na prvi par karakteristika, odnos 3:1 se održava u drugom paru karakteristika koje se odnose na oblik semena.
Mendelov drugi i treći zakon tako demonstriraju diskretnost nasljednih jedinica – jedno od najvažnijih svojstava nasljeđa.
4. ZAKON “ČISTOSTI” GAMETA.
Svaka gameta iz para alela sadrži samo jedan alel u "čistom" obliku.
MITOZIS- glavna metoda diobe ćelija sa genetski identičnom distribucijom nasljednog materijala.
U genetskom smislu Mitoza je glavni mehanizam distribucije nasljednog materijala tokom aseksualne reprodukcije organizama.
Genetski suština mitoze kao mehanizam distribucije je da se nasljedni materijal distribuira jednako u količini i kvalitetu. Posljedica toga je formiranje genetski identičnih ćelija (organizama) i nasljeđivanje karakteristika po majčinoj liniji.
MUTAGENI FAKTORI (mutageni)- faktori različite prirode čiji uticaj izaziva pojavu mutacija.
Na primjer, fizički faktori (radioaktivni zraci), hemijska jedinjenja (benzopiren i druga jedinjenja duvanskog dima), kao i delovanje virusa mogu imati mutageno dejstvo.
MUTANT- organizam u kojem je, kao rezultat mutacija, došlo do promjene jedne ili više karakteristika i svojstava.
MUTATION- nagla nasljedna promjena fenotipa uzrokovana promjenom strukture genotipa. Vidi genske, hromozomske, genomske, generativne i somatske mutacije.
MUTATIONS ALELIC- mutacije gena koje su rezultat promjena u strukturi istog gena.
GENERATIVNE MUTacije - mutacije koje nastaju kao rezultat promjena u nasljednom materijalu generativnih ćelija (gamete).
Generativne mutacije se javljaju, za razliku od somatskih, kod jedinki prve generacije potomaka (ako je mutacija dominantna) ili jedinki druge i narednih generacija u procesu homozigotizacije tokom inbridinga (ako je mutacija recesivna).
GENE MUTATIONS- mutacije koje su rezultat promjena u strukturi i funkciji jednog ili više gena.
Drugim riječima, mutacije gena nastaju na temelju promjena u strukturi molekula DNK ili RNK zbog zamjene, umetanja, gubitka nukleotida ili drugih molekularnih procesa.
SOMATSKE MUTACIJE- mutacije koje nastaju u somatskim ćelijama tijela kao rezultat promjena u njihovom nasljednom materijalu. Somatske mutacije se manifestiraju u fenotipu organizma u čijim su stanicama nastale i ne prenose se na potomke tokom spolnog razmnožavanja. Ove mutacije mogu se prenijeti na potomstvo samo aseksualnom reprodukcijom.
MUTATION SPONTANEOUS- mutacija koja je nastala pod uticajem prirodnih mutagenih faktora sredine, bez posebnih, ciljanih ljudskih akcija.
Spontane mutacije imaju važan opći biološki značaj kao materijal za prirodnu selekciju.
HROMOSOMSKE MUTACIJE- mutacije koje su rezultat preraspodjele (rekombinacije) nasljednog materijala unutar hromozoma.
Kromosomska preuređivanja (hromozomske aberacije) nastaju na osnovu jednog ili više prekida u tijelu hromozoma. Kromosomske mutacije se dijele na sljedeće vrste:
nedostatak- gubitak dijela hromozoma, inverzija- rotiranje presečenog dela hromozoma za 180° i umetanje na prvobitno mesto, umnožavanje- višestruko ponavljanje istog odsječka hromozoma, translokacija- kretanje hromozomskog segmenta unutra
jedan te isti hromozom u drugi pri razmjeni segmenata između homolognih ili nehomolognih hromozoma.
MUTAGENEZA- proces nastanka naslednih promena (mutacija) pod uticajem spoljašnjih ili unutrašnjih mutagenih faktora.
NORMA REAKCIJE- ovo je genotipski određena sposobnost organizma da, kada se promijene uslovi okoline, varira stepen izraženosti osobine u određenim granicama.
NUKLEOTID- monomer molekula nukleinske kiseline (DNK ili RNK). Nukleotidi koji čine molekule nukleinske kiseline su kombinacija ostatka fosforne kiseline, šećera (riboza ili deoksiriboza) i azotne baze (adenin, gvanin, citozin, timin ili uracil).
ONTOGENEZA- skup uzastopnih razvojnih procesa koji određuju biohemijske, fiziološke i morfološke promjene kod pojedinca od trenutka začeća do nastupanja prirodne smrti.
GLOĐENJE- spajanje muških i ženskih gameta i spajanje njihovih jezgara.
PARTENOGENEZA- djevičanska reprodukcija zasnovana na razvoju embrija iz neoplođenog jajeta.
FLOOR- skup karakteristika i svojstava organizma koji osiguravaju njegovo sudjelovanje u reprodukciji potomstva i prijenosu nasljednih informacija putem gameta, a njihovi predstavnici proizvode ženske ili muške spolne stanice - postoje jaja ili spermatozoida homogametnih i heterogametnih spolova. Kada su spolno-somatske ćelije pojedinca homogametne, polni hromozomi su predstavljeni parovima hromozoma sličnih jedni drugima (na primjer, XX).
Kod homogametnih pojedinaca, svaka gameta će sadržavati X polni hromozom. Homogametna je često žena, a heterogametna muška. Kod heterogametnog pola u somatskim ćelijama pojedinca, polni hromozomi su predstavljeni parovima različitih hromozoma (na primer, XV) ili jednim polnim hromozomom (XO), polovina gameta će imati X hromozom imaće Y hromozom (ili bez polnog hromozoma).
POLIMERIZAM- takva interakcija gena kada je razvoj neke osobine određen interakcijom dva ili više parova gena koji imaju sličan učinak na razvoj ove osobine.
Drugim riječima, polimerizacija je interakcija nealelnih gena koji jasno utiču na razvoj bilo koje osobine.
Postoje kumulativni (zbirni) i nekomulativni polimeri. Sa kumulativnom polimerizacijom, primećuje se efekat doze gena što je više dominantnih alela, gena koji deluju na polimere u genotipu, to je osobina intenzivnije izražena i obrnuto.
Kod nekomulativne polimerizacije nema efekta doziranja gena. Prisustvo jednog dominantnog alela bilo kojeg od gena koji stupaju u interakciju s polimerom dovoljno je da se osobina u potpunosti izrazi.
STANOVNIŠTVO- zbirka jedinki iste vrste, koju karakteriše zajedničko stanište i uža adaptacija na specifične uslove života -oplodne jedinke i populacije ukrštanja karakterišu različita genetska svojstva. Samooprašivači se odlikuju svojstvom homozigotizacije. Zaključak: u svakoj sljedećoj generaciji heterozigoti se smanjuju za polovicu, a broj homozigota se povećava. Suština ovog svojstva izražena je u Hardy-Weinbergovom zakonu.
U panmiktičkoj populaciji, omjer alela bilo kojeg gena ostaje isti iz generacije u generaciju.
BREED- umjetno stvorena populacija životinja koja ima određene nasljedne karakteristike. ZNAK je rezultat interakcije nasljednih informacija sa okolišnim uslovima organizma.
RODNI ZNACI- karakteristike koje su određene genima lokalizovanim na polnim hromozomima.
Postoje X-povezane i Y-povezane osobine. Svaku od ovih grupa osobina karakterišu specifične karakteristike nasleđivanja. Osobine vezane za X karakteriziraju unakrsno nasljeđivanje, dok se osobine povezane s Y karakteriziraju nasljeđivanjem po muškoj liniji (od oca na sinove, od sinova do muških unuka, itd.).
REPRODUKCIJA- svojstvo živih organizama da reprodukuju svoju vrstu, osiguravajući kontinuitet i kontinuitet života.
Aseksualno razmnožavanje- reprodukcija organizama tjelesnim stanicama bez stvaranja specijaliziranih stanica - gameta.
Postoje različite metode aseksualne reprodukcije: dioba, spore, pupanje, vegetativno.
SEKSUALNA REPRODUKCIJA je reprodukcija organizama koja se odvija kroz specijalizirane ćelije gameta.
Postoje različite metode seksualne reprodukcije: kopulacija, konjugacija, partenogeneza (apomiksis), ginogeneza, androgeneza.
RECESIVNI ALEL GENA- alel čije je djelovanje potisnuto djelovanjem drugog dominantnog alela istog gena.
RECESIVNA je osobina čiji je razvoj kod hibridne jedinke potisnut razvojem dominantne osobine.
HROMATIDI- jedna od dvije longitudinalne funkcionalne polovice hromozoma.
IZBOR- nauka o usmjerenoj transformaciji životinja, biljaka i mikroorganizama.
Svrha uzgoja je razvijanje teorije i metoda stvaranja novih rasa životinja, biljnih sorti i sojeva mikroorganizama. Selekcija se također smatra umjetnošću, jer metode selekcije stvaraju ne samo materijalne, već i duhovne vrijednosti. Tako je, na primjer, stvaranje ukrasnih sorti biljaka i ukrasnih životinja (ribe, ptice, psi) stvaralački rad, kao i rad umjetnika, muzičara itd. ::.. ima duhovnu, a ne materijalnu vrijednost.
GRADE- umjetno stvorena populacija biljaka koja ima određene nasljedne karakteristike.
TRANSKRIPCIJA- sinteza RNK molekula na odgovarajućim dijelovima DNK.
TRANSLOCATION- hromozomska mutacija koja nastaje kao rezultat razmjene sekcija između nehomolognih hromozoma. PREVOD - sinteza polipeptidnih lanaca proteina prema matrici RNK glasnika prema genetskom kodu.
FENOTIP- skup spoljašnjih i unutrašnjih karakteristika i svojstava organizma, determinisanih interakcijom genotipa sa određenim uslovima sredine.
U praksi, fenotip je rezultat interakcije genotipa sa faktorima sredine. Pojam fenotip se često koristi u pojednostavljenom smislu kada je u pitanju ispoljavanje jedne određene osobine. Na primjer: kada se ispoljava dominantna osobina - dominantni fenotip, a kada se ispoljava recesivna osobina - recesivni fenotip.
HROMOSOMSKA MAPA- grafički prikaz hromozoma koji ukazuje na udaljenost i redosled lokacije gena lokalizovanih u njemu.
HROMOSOMI- samoreproducirajuće organele ćelijskog jezgra, koje su nosioci gena koji određuju nasljedna svojstva ćelija i organizama.
CENTROMERE- mehanički centar hromozoma za koji su pričvršćene niti vretena u metafazi i anafazi mitoze i mejoze.
CITOPLAZMIČKO NASLJEĐIVANJE- nasljeđivanje osobine zbog citoplazmatskih organela koje sadrže DNK (mitohondrije, plastide, centriole).
Citoplazmatsko nasljeđivanje karakterizira nasljeđivanje po majci, jer ženske gamete sadrže veću količinu citoplazme, dok su muške gamete u pravilu lišene iste.
STRAIN- umjetno stvorena ljudska populacija mikroorganizama koja ima određene nasljedne karakteristike.
SINDROM ŠEREŠEVSKOG - TURNER- nasljedna bolest uzrokovana odsustvom jednog spolnog hromozoma. Umjesto para polnih hromozoma, u kariotipu pacijenata postoji samo jedan X hromozom. Rod je formiran kao ženski, sa karakterističnim odstupanjima od norme: primarne i sekundarne ženske polne karakteristike su nedovoljno razvijene, u pravilu takve žene
sterilni (nisu sposobni za spolno razmnožavanje), niskog su rasta, kratkog trapeznog vrata na leđnoj strani, moguća ćelavost muškog oblika, rast dlaka na prsima, kao i rast brkova i brade.
EPISTASE- ovo je interakcija između dva nealelna gena, u kojoj je djelovanje jednog gena potisnuto drugim.
Postoje dominantna i recesivna epistaza.
Dominantna epistaza je kada dominantni alel jednog gena potiskuje učinak dominantnog i recesivnog alela drugog gena.
Recesivna epistaza je kada recesivni alel jednog gena potiskuje učinak dominantnog i recesivnog alela drugog gena.
Uz populaciju u genetici postoji koncept “čista linija” je potomstvo dobijeno od samo jednog roditelja i ima potpunu sličnost s njim u genotipu
Čiste linije mogu se stvoriti u biljnoj proizvodnji od samooprašujućih biljaka. Za razliku od populacija, karakteriše ih potpuna homozigotnost. Zbog potpune homozigotnosti, selekcija u čistoj liniji je nemoguća, jer sve jedinke uključene u nju imaju identičan skup gena. Visoko homozigotni linearni miševi, pacovi i druge laboratorijske životinje stvoreni su u svrhu provođenja raznih eksperimenata, na primjer, za ispitivanje mutagenosti određenih lijekova, evaluaciju vakcina itd.
Populaciju čine životinje različitih genotipova. Efikasnost selekcije u njemu zavisi od stepena genetske varijabilnosti - omjera dominantnih i recesivnih gena. Hardy i Weinberg su proveli matematičku analizu distribucije gena u velikim populacijama u kojima nema selekcije, mutacije ili miješanja populacija. Utvrdili su da je takva populacija u stanju ravnoteže u smislu omjera genotipova, koji je određen formulom
r2AA + + IpqAa + q)aa =1,
gdje je p frekvencija dominantnog gena A,
q je frekvencija njegovog recesivnog alela a.
U skladu s tim, formuliran je Hardy-Weinbergov zakon, odnosno pravilo, prema kojem, u nedostatku faktora koji mijenjaju frekvencije gena, populacije, u bilo kojem odnosu alela iz generacije u generaciju, održavaju ove frekvencije alela konstantnim. Uprkos poznatim ograničenjima, koristeći Hardy-Weinbergovu formulu, moguće je izračunati strukturu populacije i odrediti frekvencije heterozigota (na primjer, za smrtonosne ili subletalne gene, poznavajući učestalosti homozigota za recesivne osobine i učestalosti jedinki sa dominantna osobina), analiziraju pomake u frekvencijama gena za specifične osobine kao rezultat selekcije, mutacija i drugih faktora.
Populacija je u ravnoteži samo kada u njoj nema selekcije. Kada se pojedinačne životinje u takvoj populaciji odstrele, mijenja se omjer gameta, što utiče na genetsku strukturu sljedeće generacije. Međutim, K. Pearson je pokazao da čim nastane stanje panmiksije (slobodnog ukrštanja), odnos genotipova i fenotipova u populaciji u sljedećoj generaciji se vraća na onaj koji odgovara Hard-Weinbergovoj formuli, ali s drugačijim odnos. Ukrštanje koje obnavlja omjer genotipova u populaciji, u skladu s Hardy-Weinberg formulom, naziva se stabilizacijom. Iz ovoga slijedi zaključak: kada se u populaciji koriste nasumični, neselektirani bici ili matice, dolazi do stabilizacije osobina produktivnosti na istom nivou, a povećanje produktivnosti životinja u takvoj situaciji je nemoguće. Slično, u nedostatku odstrela heterozigotnih nosilaca recesivnih anomalija, učestalost abnormalnih životinja u populaciji ostaje nepromijenjena.
U populacijama domaćih životinja frekvencije gena se konstantno mijenjaju, što se može uočiti pri analizi susjednih generacija. Takve promjene su suština genetske evolucije. Glavni faktori evolucije: mutacije, prirodna i umjetna selekcija, migracija, genetski drift.
Jedan od glavnih uzroka genetskih varijacija u populaciji su mutacije. Spontane mutacije svakog gena javljaju se na niskoj frekvenciji, ali je ukupna učestalost mutacija svih gena koje sadrže populacije vrlo visoka. Mutacije koje se javljaju u zametnim stanicama roditeljske generacije dovode do promjena u genetskoj strukturi potomstva. U populaciji stalne veličine, u nedostatku selekcije, većina mutacija koje nastaju brzo se gube, ali neke od njih mogu opstati kroz nekoliko generacija. Nestanak mutantnih gena iz populacije suprotstavlja se djelovanjem procesa mutacije, što rezultira stvaranjem ponovljenih mutacija.
Genetska struktura populacija se formira i mijenja pod utjecajem prirodne i umjetne selekcije. Efekat prirodne selekcije je da jedinke sa visokom održivošću, ranom zrelošću, plodnošću itd., odnosno prilagođenije uslovima sredine, imaju preferencijalnu reprodukciju. U vještačkoj selekciji, osobine produktivnosti su od odlučujućeg značaja.
IN AND. Vlasov napominje da se prirodna selekcija javlja u svim fazama ontogeneze populacije - od formiranja gameta do odraslog organizma. Istovremeno, značajno utiče na brzinu veštačke selekcije zbog suprotnog efekta tokom selekcije za visok nivo razvoja produktivnih osobina, što je neuobičajeno za biološke granice vrste. Na osnovu toga, prilikom odabira životinja potrebno je uzeti u obzir ne samo proizvodne osobine, već i znakove prilagodljivosti uvjetima okoline.
Prema S.M. Geršenzona, kriterij intenziteta prirodne selekcije je razlika u sposobnosti upoređenih grupa, nazvana selekcijski koeficijent i izražena u dijelovima jedinice. Na primjer, ako je vjerovatnoća da će jedinke sa genotipom aa ostaviti potomstvo 10% manja nego kod individua sa AA ili Aa genotipom, tada je sposobnost ovih grupa za jedinke AA i Aa jednaka 1, za jedinke aa - 0,9 .
Sa stanovišta veterinarske genetike, važna je efikasnost selekcije protiv štetnih mutacija, posebno recesivnog tipa. Analiza pokazuje da se visoke frekvencije recesivnog mutantnog gena mogu brzo smanjiti na niske vrijednosti selekcijom. Za smanjenje učestalosti smrtonosnog gena, na primjer sa 0,3 na 0,2, dovoljne su dvije generacije.
Učestalost homozigota (aa) za mutantni gen ovisi o učestalosti heterozigotnih životinja u populaciji. Identifikacija ovih heterozigota i njihovo eliminisanje u skladu s tim će smanjiti učestalost genetskih abnormalnosti uzrokovanih mutiranim genom, što je posebno važno kada je učestalost mutacija visoka.
Genetska struktura populacije može se promijeniti zbog slučajnih genetsko-automatskih procesa (prema N.P. Dubininu) ili genetskog drifta (prema S. Wrightu). Zapažanja pokazuju da se genetski drift najintenzivnije javlja u malim populacijama. Na primjer, poznati su slučajevi visokih koncentracija rijetkih mutacija u malim izolovanim populacijama goveda i drugih životinjskih vrsta, očigledno povezanih s genetsko-automatskim procesima. Širenje mutacija u različitim životinjskim populacijama može nastati kao rezultat migracija.
Parenje blisko srodnih životinja naziva se inbreeding. Parenje u srodstvu, ili inbreeding, je metoda selekcije koja se koristi u stočarstvu za konsolidaciju vrijednih nasljednih osobina određene životinje u narednim generacijama. Životinje koje su povezane jedna s drugom pokazuju sličnosti u određenim parovima alela koje su primile od zajedničkog pretka. Što je bliži stepen veze, to je veća sličnost.
Svaka životinja u svom genotipu ima alelne gene, kako u homozigotnom tako iu heterozigotnom stanju. Heterozigot obično sadrži štetne mutantne recesivne gene. Sa inbreedingom, povećava se vjerovatnoća fuzije identičnih gameta koje nose mutantne gene u heterozigotnom stanju i njihovog prijelaza u homozigotno stanje. Ova vjerovatnoća je proporcionalna stepenu srodstva sparenih životinja.
Dakle, kao rezultat korištenja inbreedinga dolazi do promjene frekvencije gena, povećava se vjerojatnost odvajanja recesivnih homozigota, što je uzrok inbreeding depresije, izražene u smanjenju vitalnosti i plodnosti životinja, te rođenju abnormalne osobe.
Inbreeding je u pravilu bio složen - istovremeno na dva (1. znamenka) ili tri (2. znamenka) predaka.
U smislu pokazatelja koji karakteriziraju produktivnost i održivost životinja, urođena depresija nije fatalna popratna pojava inbreedinga.
Mnogo je primjera u kojima nisu uočene negativne posljedice prilikom inbridinga različitog stepena, uključujući i bliske.
N.P. S tim u vezi, Dubinin napominje da „linija propada dok prolazi kroz procese sekvencijalne akumulacije štetnih recesivnih gena koji prelaze u homozigotno stanje. Kada dođe do manje ili više izraženog završetka ovog procesa, linije postaju relativno konstantne u svojim svojstvima i mogu ostati u tako stabilnom stanju dugo vremena. Samo nove mutacije koje se akumuliraju u njima mogu promijeniti genotip takvih linija.” Međutim, naglašava akademik, “mnoge linije, naravno, umiru tokom inbridinga, jer u njima smrtonosni i poluletalni geni prelaze u homozigotno stanje”. Stoga se inbreeding koristi kao metoda individualne selekcije za prijenos vrijednih gena izvanrednih životinja u homozigotno stanje.
Tokom duge evolucije životinja, zajedno sa korisnim mutacijama prikupljenim selekcijom, u populacijama ili rasama nakupio se određeni spektar genskih i hromozomskih mutacija. Svaka generacija populacije nasljeđuje ovo opterećenje mutacija i u svakoj od njih nastaju nove mutacije, od kojih se neke prenose na sljedeće generacije.
Očigledno je da se većina štetnih mutacija odbacuje prirodnom selekcijom ili eliminiše tokom procesa selekcije. To su prvenstveno dominantne genske mutacije, fenotipski manifestirane u heterozigotnom stanju, i kvantitativne promjene u hromozomskim setovima. Kroz selekcijsko sito mogu proći recesivno djelujuće genske mutacije u heterozigotnom stanju i strukturna preuređivanja hromozoma, koji ne utiču značajno na održivost njihovih nosilaca. Oni čine genetski teret populacije. Dakle, genetsko opterećenje populacije shvata se kao skup štetnih genskih i hromozomskih mutacija. Pravi se razlika između mutacionog i segregacionog genetskog opterećenja. Prvi nastaje kao rezultat novih mutacija, drugi - kao rezultat cijepanja i rekombinacije alela pri križanju heterozigotnih nositelja "starih" mutacija.
Učestalost smrtonosnih, poluletalnih i subvitalnih mutantnih gena koji se prenose s generacije na generaciju u obliku mutacijskog genetskog opterećenja ne može se precizno izbrojati zbog poteškoća u identifikaciji nosilaca. Morton i Crowe predložili su obrazac za izračunavanje nivoa genetskog opterećenja u broju smrtonosnih ekvivalenata. Jedan smrtonosni ekvivalent odgovara jednom smrtonosnom genu koji uzrokuje smrtnost s vjerovatnoćom od 10%, dva smrtonosna gena sa vjerovatnoćom smrti od 50%, itd. Vrijednost genetskog opterećenja prema Mortonovoj formuli:
log eS = A + BF,
gdje je S dio potomstva koji ostaje živ;
A - mortalitet, mjeren smrtonosnim ekvivalentom u populaciji pod uslovom slučajnog parenja (F = 0), plus mortalitet zbog vanjskih faktora;
B je očekivano povećanje mortaliteta kada populacija postane potpuno homozigotna (F = 1);
F - koeficijent inbridinga.
Nivo genetskog opterećenja može se odrediti na osnovu fenotipske manifestacije mutacija (deformiteti, urođene metaboličke abnormalnosti, itd.), analize vrste njihovog naslijeđa i učestalosti u populaciji.
N.P. Dubinin predlaže da se odredi genetsko opterećenje populacije upoređivanjem učestalosti mrtvorođenih u srodnim i nepovezanim selekcijama roditeljskih parova. Treba imati na umu da uz visoku učestalost heterozigota za recesivne smrtonosne i poluletalne mutantne gene, rođenje životinja s anomalijama ne mora nužno biti povezano sa bliskim i umjerenim stupnjevima inbreedinga. Zajednički predak (izvor mutacije) također se može nalaziti u udaljenim redovima pedigrea. Na primjer, bik Truvor 2918, heterozigotni nosilac mutantnog recesivnog gena, bio je u V, VI, VII redovima predaka na državnoj farmi Crvena Baltika, ali kada je njegov pra-praunuk Avtomat 1597 korišten na srodnim kravama , uočeni su masovni slučajevi rođenja golih teladi.
Ovi podaci u određenoj mjeri karakterišu nivoe genetskog opterećenja pojedinih mutantnih gena u specifičnim populacijama goveda.
Kromosomske mutacije su dio genetskog opterećenja. Snimaju se direktnom citološkom metodom. Prema rezultatima brojnih studija, glavna komponenta opterećenja hromozomskih aberacija kod goveda su Robertsonove translokacije, a kod svinja - recipročne. Najčešća mutacija kod goveda bila je translokacija hromozoma 1/29. Raspon varijabilnosti učestalosti ove aberacije, prema našim podacima, u populacijama bledo obojenih goveda kretao se od 5 do 26%.
Dakle, koncept genetskog opterećenja u svjetlu modernog napretka citogenetike treba proširiti. Sada kada je poznat širok spektar hromozomskih aberacija i utvrđeno striktno nasljeđivanje pojedinih od njih (translokacije i inverzije), čini se preporučljivim uzeti ih u obzir uz štetne genske mutacije kao dio genetskog opterećenja.
Postojanje nasledne varijabilnosti u populacijama, prvenstveno mutacija u heterozigotnom stanju, omogućava im da se brzo prilagode novim uslovima sredine promenom genetske strukture. Proces mutacije također dovodi do formiranja genetskog polimorfizma u populacijama – raznolikost u frekvencijama alela, homozigoti za dominantne gene, heterozigoti ili homozigoti za recesivne gene. Polimorfizam je mehanizam koji održava postojanje populacija. Ako, na primjer, heterozigotnost osigurava bolju prilagodljivost promijenjenim uvjetima okoline, tada dolazi do selekcije u korist heterozigota, što dovodi do uravnoteženog polimorfizma – reprodukcije u populaciji iz generacije u generaciju određenog omjera različitih genotipova i fenotipova. Procesi koji osiguravaju sposobnost populacije da održi svoju genetsku strukturu nazivaju se genetska homeostaza.
U genetici postoje dvije klase osobina - kvalitativne i kvantitativne. Razlikuju se po prirodi varijabilnosti i karakteristikama nasljeđivanja. Kvalitativne karakteristike karakteriše povremena, a kvantitativne - kontinuirana varijabilnost. Prvi od njih daju jasne granice kada se dijele na dominantne ili recesivne osobine. To je zato što svaki od njih obično kontrolira jedan alelni gen. Kvantitativne osobine ne daju jasne granice za razdvajanje pod različitim opcijama ukrštanja, iako se razlikuju od kvalitativnih osobina u većem stepenu varijabilnosti. Karakteristika kvantitativnih osobina je složena priroda nasljeđivanja. Svaki od njih nije određen jednim, već mnogim lokusima u hromozomima. Ova vrsta nasljeđivanja, kada je jedna osobina određena mnogim genima, naziva se poligenom. Nivo razvoja kvantitativne osobine zavisi od odnosa dominantnih i recesivnih gena, drugih genetskih faktora i stepena modifikacionog dejstva faktora sredine. Varijabilnost kvantitativne osobine u populaciji sastoji se od genetske i paraloške (eksterne sredine) varijabilnosti.
Pojam heritabilnosti osobina i koeficijenta heritabilnosti. Različite kvantitativne osobine imaju različite stepene genetske varijabilnosti, a uslovi sredine imaju različite efekte na nivo fenotipske manifestacije određene osobine. Prilikom odabira životinja od najveće je važnosti znati u kojoj mjeri će postojati podudarnost u razinama razvoja kvantitativne osobine kod roditelja i potomstva, odnosno u kojoj će mjeri potomstvo naslijediti kvantitativne ekonomski korisne osobine ili patološke osobine roditelji.
Ekonomski korisna svojstva obuhvataju proizvodnju mlijeka, sadržaj masti i bjelančevina u mlijeku krava, šišanje vune kod ovaca, proizvodnju jaja kod pilića, prirast žive mase, plodnost itd. Povećanje stepena razvijenosti ekonomski korisnih osobina postiže se stalnim odabirom najbolje jedinke za reprodukciju. Efikasnost selekcije za ove osobine zavisi od stepena njihove heritabilnosti, odnosa između njih, razlike između prosečne vrednosti osobine odabrane grupe i proseka za stado (selekcijski diferencijal) i intervala između generacija.
Vrijednosti koeficijenata heritabilnosti zavise od prirode osobine. Tako je prosečna vrednost A2 za mlečnost 0,25, sadržaj mlečne masti 0,38, živa masa kod ovaca je 0,35, prinos čiste vune je 0,55, plodnost kod goveda je 0,08 itd.
Selekciona praksa i posebna istraživanja akumulirali su podatke koji ukazuju da u nekim slučajevima stepen razvijenosti jedne ili više osobina u potomstvu premašuje stepen izraženosti ovih osobina kod najboljih roditelja. Ovaj fenomen, nazvan heterozis, ne uklapa se u uobičajeni okvir nasljeđivanja osobina. Predložene su različite hipoteze da se to objasni:
1) heterozigotno stanje za mnoge gene;
2) interakcije dominantnih povoljnih gena;
3) overdominacija, kada su heterozigoti superiorni u odnosu na homozigote.
N.V. Turbin je predložio teoriju genetske ravnoteže, koja se zasniva na složenoj prirodi uzročno-posledičnih veza između naslednih faktora i osobina.
N.G. Dmitriev i I.JI. Halperin napominje da se glavni razlog za nastanak heterozisa mora tražiti u posebnostima evolucije vrste, pasmine, linije. Treba imati na umu da je sve u prirodi usmjereno na očuvanje života. Manifestacija heteroze zavisi od genetske prirode osobine. Dakle, prilikom ukrštanja ili međulinijskog ukrštanja, heteroza je izraženija u odnosu na osobine koje imaju nizak stepen heritabilnosti.
Što se tiče osobina koje imaju prosječnu ili visoku heritabilnost, heteroza se kod njih najčešće slabo manifestira, a hibridi obično zauzimaju srednji položaj.
U prisustvu prave heteroze, vrijednost indeksa je ili veća od 100%. Ako je vrijednost heterozisa manja od 100% ili ima predznak minus, tada je ispravnije govoriti o boljoj ili lošijoj kombinacijskoj sposobnosti linija. Ukrštanje potonjeg po određenom obrascu osigurava kod hibridnog potomstva najbolji razvoj jedne osobine od oca, a druge od majke, iako ova osobina u hibridu u svom razvoju ne nadmašuje najbolji roditeljski oblik.